![]()
研究プロジェクトⅡ
幹細胞アーキオサイト、分化細胞を遺伝子発現で捉える
私達が研究を始めた2002年には、カイメン研究において分子生物学的手法がまだ確立していず、カイメンの細胞種については細胞形態によって細胞種を識別している状態でした。
例えばアーキオサイトは、「大型で、活発に分裂し、活発に移動し、大きな1つの核小体を含む大型の核を持つ細胞。芽球からの個体発生の初期には栄養顆粒であるビテリン小片を多数もつ」という下図左・中央にあるような形態の細胞が、幹細胞アーキオサイトとして同定されていました。しかし、全ての細胞種が幹細胞アーキオサイトから分化するのですから、似た細胞形態の細胞種もあるはずですし、分化途中のアーキオサイトもあるはずです。
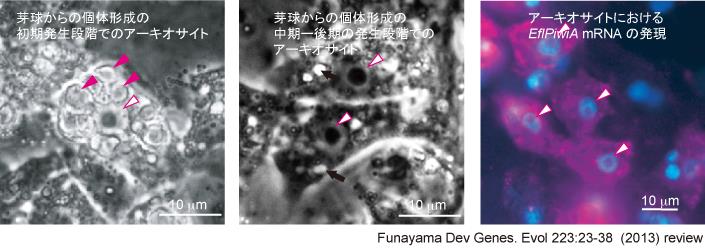
どれが幹細胞アーキオサイトで、どれが分化途中のアーキオサイト、どれが特定の分化細胞種なのかをきちんと遺伝子発現で同定出来る、遺伝子解析法をカイメンで確立すること、1細胞レベルで識別可能な高解像な実験系を確立することが私達の第一歩でした。
私達は、独自にカワカイメンESTライブラリーの作成(網羅的な遺伝子配列を得る)1細胞レベルで検出可能な高感度のmRNA発現の検出(in situ hybridization法)、抗体染色法など基本的な手法を確立し、ごく最近では、工夫を凝らして細胞種特異的なトランスクリプトーム解析なども行っています。
| これまでに複数種類の分化細胞に関して細胞系列特異的な遺伝子を同定、遺伝子発現による細胞種の同定を可能にしてきました。例えば襟細胞が食細胞である事を生かした工夫を考え、蛍光ビーズを非常に短時間与えることで、下図の様に襟細胞のみをラベル、その後細胞を解離しFACSにかけることで、取り込んだ蛍光ビーズの蛍光がある小型の細胞として分取、プロテオーム(網羅的な発現タンパク質解析)から、襟細胞系列特異的に発現する遺伝子を同定しています。 中でも、動植物の幹細胞で発現し短い特定のRNA(piRNA)を介して幹細胞性の維持に働くと知られているPiwiのホモログ(EflPiwiA, EflPiwiB)が、カイメン幹細胞アーキオサイトでも発現し、分化過程でEflPiwiA 及び EflPiwiB mRNAの発現が失われることを明らかにしました。Piwiを介した幹細胞性維持機構は、カイメンにも存在し、これは多細胞動物の起源的な幹細胞分子基盤の1つであることが明らかになりました。ゲノム配列の読まれたおなじく普通カイメンに属するAmphymedon queenslandicaで、piRNAが複数報告されていることも、piRNAを介したPIWIによる幹細胞維持機構がカイメンにあることを支持しています。 |
 |
 |
襟細胞はEflPiwiA, EflPiwiBの発現を維持しており、幹細胞性が維持されているらしい
EflPiwiAの発現解析の実験から、さらに興味深いことが示唆されました。上述のように、分化細胞ではEflPiwiAの発現は失われるのですが、唯一の例外として、「襟細胞」ではずっとEflPiwiAの発現が維持させていたのです。
襟細胞は1本の鞭毛とそれを囲む微絨毛が側面で結合した筒状の襟を持つ、という特徴的な細胞形態を持ち、さらに一層に内向きに並んで袋状上皮構造を作り、水管の側面に繋がっていて、i)鞭毛を動かすことで水管の中に流れを生じさせる、ii)カイメンの体の外から水管に入ってきた微小な栄養物を食し栄養を他の細胞に顆粒として受け渡す、という特定の機能を持つ分化細胞です。
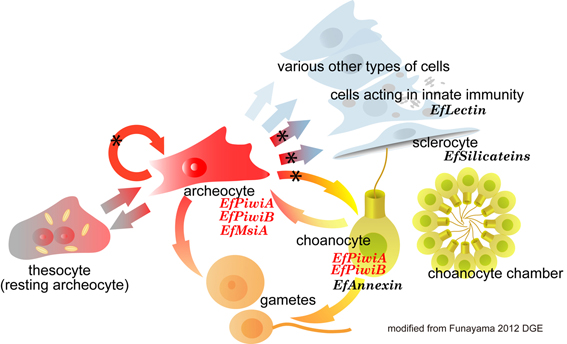
ふつうカイメン綱の幹細胞システムは、アーキオサイトと襟細胞の2細胞種から成る
成熟した襟細胞でEflPiwiAが発現することは予想外でしたが、考えてみると、カイメンは生殖幹細胞というものが無く、有性生殖時に配偶子を生み出すのは、アーキオサイトと襟細胞です。種によって違いますが、ふつうカイメン綱では通常アーキオサイトから卵が、襟細胞から精子が減数分裂により生み出されます。また、非常に古い組織学的な解析の論文ですが、再生時や芽球形成時に襟細胞が脱上皮化してアーキオサイトへと変化するという報告もあります。これらのことを考え合わせると、襟細胞は通常は食細胞として働きながら、有性生殖時に配偶子を形成する時、また、多数のアーキオサイトが必要となる芽球形成や再生時に、アーキオサイトへと変化する、いわばストック型幹細胞として働いているのだろうと考えられます。
私達は普通カイメン綱のカイメン幹細胞システムは、アーキオサイトと襟細胞という2種類の細胞により構成され、無性生殖、有性生殖、個体の恒常性の維持を担っているというモデルを提唱しています。
特異的に発現するEflMusashiAの同定と詳細な解析からみえてきた分裂様式
EflPiwiA, EflPiwiBを発現する大型の細胞としてアーキオサイトを同定出来るようになったことで解析が進み、アーキオサイトでのみ特異的に発現する遺伝子EflMsiAを同定しました。Musashiはもともとショウジョウバエの発生において、神経幹細胞の維持に必要な遺伝子として同定され、最近ではES細胞やその他の動物の組織幹細胞でも特異的な発現があることが知られるようになっている遺伝子です。特異的な抗体が作成出来たので、細胞内局在を見たところ、興味深いことに、EflMsiAタンパク質は、細胞質ばかりでなく、核内に強く局在していることが分かりました。最近になってESや他の動物の組織幹細胞でもMusashiタンパク質の核内の局在が報告され、核内でmiRNA合成系を調節し、遺伝子発現制御を行っているという報告もあります。カイメンでアーキオサイト特異的に発現していることから、起源的な幹細胞分子基盤の一つとして、Musashiタンパク質による遺伝子発現制御機構が示唆されました。
幹細胞の調節機構の一つに、分化細胞を生み出しながらも幹細胞集団を枯渇させない機構が考えられます。個体形成過程において、生み出す分化細胞の数の制御が必要な場合に不等分裂が用いられている例は古くから研究されてきました。では、カイメンにおいてはどの様に分化細胞が生み出されているのでしょうか?
この疑問に答えるために、分裂期アーキオサイトにおけるEflMsiA mRNA・タンパク質の発現と局在の詳細な解析を行い、i)分裂期アーキオサイトには、EflMsiA mRNA・タンパク質の偏りはみられず、1方の娘細胞のみに受け渡されるという不等分裂は示唆されない、ii)EflMsiA mRNA・タンパク質の量に差のある2つのタイプに分けられる、という結果から、アーキオサイトは当分裂を行っていることが示唆されました。
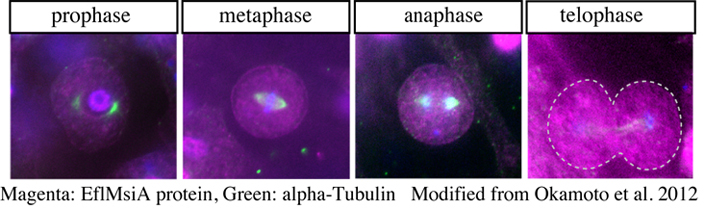
| 芽球からの個体形成においてカイメンの体は急速に成長していることを考えると、それぞれの分化細胞を生み出す割合と、幹細胞の自己複製の足とが別々に制御できる等分裂は、状況にあった優れた仕組みだと考えられます。ヒドラ、プラナリアなど体制多能性幹細胞を持つ動物において、幹細胞の分裂様式を明らかにした例は他になく、今後これらの動物での多能性幹細胞の分化・自己複製の制御機構の理解の1つの参考にもなると考えています。 | 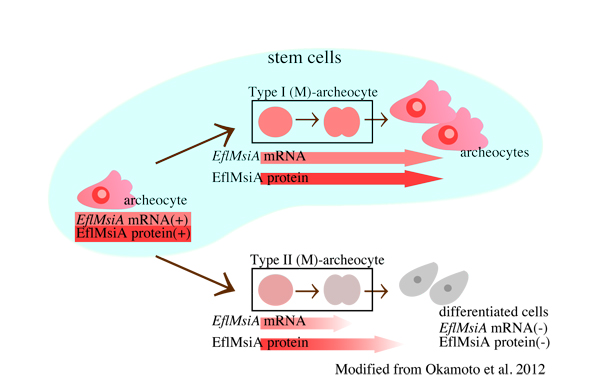 |
※画像クリックで拡大します。



本サイトの掲載の文章・写真の無断転載を禁じます。